
33 Muscular functions and anatomy
Learning Objectives
After reading this section, you should be able to-
- Describe the major functions of muscle tissue.
- Describe the components within a skeletal muscle fiber (e.g., sarcolemma, transverse [T] tubules, sarcoplasmic reticulum, myofibrils, thick [myosin] myofilaments, thin [actin] myofilaments, troponin, tropomyosin).
- Define sarcomere.
- Describe the arrangement and composition of the following components of a sarcomere: A-band, I-band, H-zone, Z-disc (line), and M-line.
- Describe the structure of the neuromuscular junction.
Muscle is one of the four primary tissue types of the body (along with epithelial, nervous, and connective tissues), and the body contains three types of muscle tissue: skeletal muscle, cardiac muscle, and smooth muscle (Figure 33.1). All three muscle tissues have some properties in common; they all exhibit a quality called excitability as their plasma membranes can change their electrical states (from polarized to depolarized) and send an electrical wave called an action potential along the entire length of the membrane. While the nervous system can influence the excitability of cardiac and smooth muscle to some degree, skeletal muscle completely depends on signaling from the nervous system to work properly. On the other hand, both cardiac muscle and smooth muscle can respond to other stimuli, such as hormones and local stimuli.
A unique property common to all three types of muscle is contractility, which is the ability of the cells to shorten and generate force. While muscle tissue can shorten with contractions, it also displays extensibility or the ability to stretch and extend beyond the resting length of the cells. After being stretched, the elasticity of muscle allows it to recoil back to its original length.
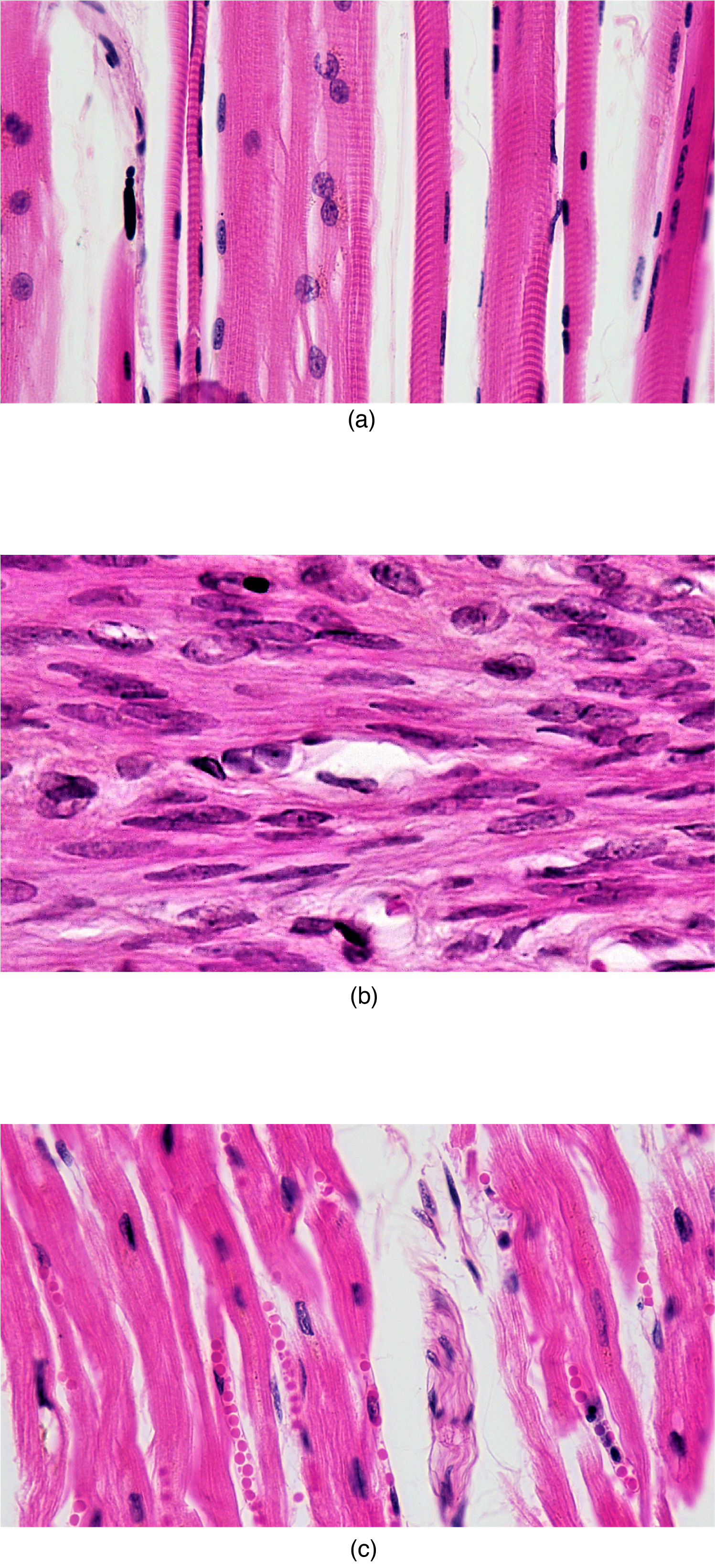
The muscles all begin the mechanical process of contracting (shortening) when a protein called actin is pulled by a protein called myosin, and differences in the microscopic organization of these contractile proteins exist among the three muscle types. In both skeletal and cardiac muscle, the actin and myosin proteins are arranged very regularly in the cytoplasm of individual muscle cells, which creates an alternating light and dark striped pattern called striations. The striations are visible with a light microscope under high magnification (see Figure 33.1). Smooth muscle (named for its lack of striations), does not produce this striped pattern because the contractile proteins are not arranged in such regular fashion.
Skeletal muscle cells (also called muscle fibers) are unique in that they are multinucleated , with the nuclei located on the periphery of the cell under the cell plasma membrane (also called sarcolemma in muscle). During early development, embryonic myoblasts, each with its own nucleus, fuse with hundreds of other myoblasts to form long multinucleated skeletal muscle fibers. Cardiac muscle cells each generally have one nucleus centrally located in the cell, but the cells are physically and electrically connected to each other so that the contraction signals spread through cells and the entire heart contracts as one unit. Smooth muscle cells contain a single nucleus and can exist in electrically linked units contracting together as a single-unit or as multi-unit smooth muscle where cells are not electrically linked.
Muscle Functions
The best-known feature of skeletal muscle is its ability to contract and cause movement. Skeletal muscles act not only to produce movement but also to stop movement, such as resisting gravity to maintain posture. Small, constant adjustments of the skeletal muscles are needed to hold a body upright or balanced in any position. Muscles also prevent excess movement of the bones and joints, maintaining skeletal stability and preventing skeletal structure damage or deformation. Skeletal muscles are located throughout the body at the openings of internal tracts to control the movement of various substances. These muscles allow functions, such as swallowing, urination, and defecation to be under voluntary control. Skeletal muscles also protect internal organs (particularly abdominal and pelvic organs) by acting as an external barrier or shield to external trauma and supporting the weight of the organs.
Skeletal muscles contribute to the maintenance of homeostasis in the body by generating heat. Muscle contraction requires energy and, when ATP is broken down, heat is produced. This heat is very noticeable during exercise, when sustained muscle movement causes body temperature to rise, and in cases of extreme cold, when shivering produces random skeletal muscle contractions to generate heat.
Cardiac muscle is only found in the heart and functions to generate force and build pressure gradients to drive blood flow throughout the body. Smooth muscle in the walls of arteries is a critical component that regulates blood pressure and blood flow through the circulatory system. Smooth muscle in the skin, visceral organs, and internal passageways is also essential for moving materials through the body. Neither cardiac nor smooth muscle connect to bone and therefore they cannot produce the gross movements we associate with skeletal muscle.
Skeletal muscle
Each skeletal muscle is an organ that consists of various integrated tissues. These tissues include the skeletal muscle fibers, blood vessels, nerve fibers, and connective tissue. Each skeletal muscle has three layers of connective tissue (called mysia) that enclose it, provide structure to the muscle, and compartmentalize the muscle fibers within the muscle (Figure 33.2). Each muscle is wrapped in a sheath of dense, irregular connective tissue called the epimysium, which allows a muscle to contract and move powerfully while maintaining its structural integrity. The epimysium also separates muscle from other tissues and organs in the area, allowing the muscle to move independently.
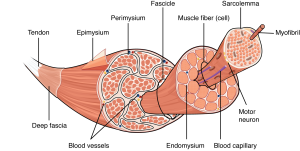
Inside each skeletal muscle, muscle fibers are organized into bundles, called fascicles, surrounded by a middle layer of connective tissue called the perimysium. This fascicular organization is common in muscles of the limbs; it allows the nervous system to trigger a specific movement of a muscle by activating a subset of muscle fibers within a fascicle of the muscle. Inside each fascicle, each muscle fiber is encased in a thin connective tissue layer of collagen and reticular fibers called the endomysium. The endomysium surrounds the extracellular matrix of the cells and plays a role in transferring force produced by the muscle fibers to the tendons.
In skeletal muscles that work with tendons to pull on bones, the collagen in the three connective tissue layers intertwines with the collagen of a tendon. At the other end of the tendon, it fuses with the periosteum coating the bone. The tension created by contraction of the muscle fibers is then transferred though the connective tissue layers, to the tendon, and then to the periosteum to pull on the bone for movement of the skeleton. In other places, the mysia may fuse with a broad, tendon-like sheet called an aponeurosis, or to fascia, the connective tissue between skin and bones. The broad sheet of connective tissue in the lower back that the latissimus dorsi muscles (the “lats”) fuse into is an example of an aponeurosis.
Every skeletal muscle is also richly supplied by blood vessels for nourishment, oxygen delivery, and waste removal. In addition, every muscle fiber in a skeletal muscle is supplied by the axon branch of a somatic motor neuron, which signals the fiber to contract. Unlike cardiac and smooth muscle, the only way to functionally contract a skeletal muscle is through signaling from the nervous system.
Skeletal Muscle Fibers
Because skeletal muscle cells are long and cylindrical, they are commonly referred to as muscle fibers (or myofibers). Skeletal muscle fibers can be quite large compared to other cells, with diameters up to 100 μm and lengths up to 30 cm (11.8 in) in the Sartorius of the upper leg. Having many nuclei allows for production of the large amounts of proteins and enzymes needed for maintaining normal function of these large protein dense cells. In addition to nuclei, skeletal muscle fibers also contain cellular organelles found in other cells, such as mitochondria and endoplasmic reticulum. However, some of these structures are specialized in muscle fibers. The specialized smooth endoplasmic reticulum, called the sarcoplasmic reticulum (SR), stores, releases, and retrieves calcium ions (Ca2+).
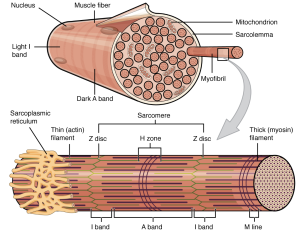
The plasma membrane of muscle fibers is called the sarcolemma (from the Greek sarco, which means “flesh”) and the cytoplasm is referred to as sarcoplasm (Figure 33.3). Within a muscle fiber, proteins are organized into organelles called myofibrils that run the length of the cell and contain sarcomeres connected in series. Because myofibrils are only approximately 1.2 μm in diameter, hundreds to thousands (each with thousands of sarcomeres) can be found inside one muscle fiber. The sarcomere is the smallest functional unit of a skeletal muscle fiber and is a highly organized arrangement of contractile, regulatory, and structural proteins. It is the shortening of these individual sarcomeres that lead to the contraction of individual skeletal muscle fibers (and ultimately the whole muscle).
The Sarcomere
A sarcomere is defined as the region of a myofibril contained between two cytoskeletal structures called Z-discs (also called Z-lines or Z-bands, Table 33.1), which anchor the thin filaments and define the boundary of each sarcomere, and the striated appearance of skeletal muscle fibers is due to the arrangement of the thick and thin myofilaments within each sarcomere (Figure 33.3). The dark striated A band is composed of the thick filaments containing myosin, which span the center of the sarcomere extending toward the Z-dics. The thick filaments are anchored at the middle of the sarcomere (the M-line) by a protein called myomesin. The lighter I band regions contain thin actin filaments anchored at the Z-discs by a protein called α-actinin. The thin filaments extend into the A band toward the M-line and overlap with regions of the thick filament. The A band is dark because of the thicker myosin filaments as well as overlap with the actin filaments. The H zone in the middle of the A band is a little lighter in color because it only contains the portion of the thick filaments that does not overlap with the thin filaments (i.e. the thin filaments do not extend into the H zone).
Because a sarcomere is defined by Z-discs, a single sarcomere contains one dark A band with half of the lighter I band on each end (Figure 33.4). During contraction the myofilaments themselves do not change length, but actually slide across each other so the distance between the Z-discs shortens resulting in the shortening of the sarcomere. The length of the A band does not change (the thick myosin filament remains a constant length), but the H zone and I band regions shrink. These regions (e.g., H-zone and I-band) represent areas without filament overlap, and as the overlap increases during contraction, the length of these non-overlapping regions decrease. The A-band stays the same. The sarcomere, H-zone, and I-band shorten during contraction.
| Component | Location | Function |
|---|---|---|
| Z-disc | Sarcomere boundary | Anchors thin filaments |
| M-line | Center of sarcomere | Anchors thick filaments |
| I-band | Contains only thin filaments | Shortens during contraction |
| A-band | Overlaps thick and thin filaments | Stays the same during contraction |
| H-zone | Only thick filaments | Shortens during contraction |
Myofilament Components
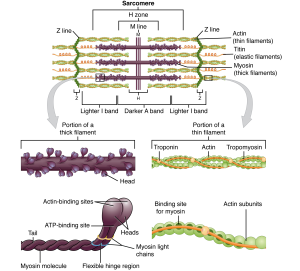
The thin filaments are composed of two filamentous actin chains (F-actin) comprised of individual actin proteins (Figure 33.4). These thin filaments are anchored at the Z-disc and extend toward the center of the sarcomere. Within the filament, each globular actin monomer (G-actin) contains a myosin binding site and is also associated with the regulatory proteins, troponin and tropomyosin. The troponin protein complex consists of three polypeptides. Troponin I (TnI) binds to actin, troponin T (TnT) binds to tropomyosin, and troponin C (TnC) binds to calcium ions. Troponin and tropomyosin run along the actin filaments and control when the actin binding sites will be exposed actin-myosin binding.
Thick myofilaments are composed of myosin protein complexes, which are composed of six proteins: two myosin heavy chains and four light chain molecules. The heavy chains consist of a tail region, flexible hinge region, and globular head which contains an actin-binding site and a binding site for the high energy molecule ATP. The light chains play a regulatory role at the hinge region, but the heavy chain head region interacts with actin and is the most important factor for generating force. Hundreds of myosin proteins are arranged into each thick filament with tails toward the M-line and heads extending toward the Z-discs.
Other structural proteins are associated with the sarcomere but do not play a direct role in active force production. Titin, which is the largest known protein, helps align the thick filament and adds an elastic element to the sarcomere. Titin is anchored at the M-line, runs the length of myosin, and extends to the Z-disc. The thin filaments also have a stabilizing protein, called nebulin, which spans the length of the thick filaments.
The Sliding Filament Model of Contraction
The arrangement and interactions between thin and thick filaments allows for the sarcomeres to generate force. When a motor neuron releases a neurotransmitter at the neuromuscular junction, a skeletal muscle fiber is activated. Cross-bridges form between the thick and thin filaments and the thin filaments are pulled which slide past the thick filaments within the fiber’s sarcomeres. It is important to note that while the sarcomere shortens, the individual proteins and filaments do not change length but simply slide next to each other. This process is known as the sliding filament model of muscle contraction (Figure 33.5).

The filament sliding process of contraction can only occur when myosin-binding sites on the actin filaments are exposed by a series of steps that begins with Ca2+ entry into the sarcoplasm. Tropomyosin winds around the chains of the actin filament and covers the myosin-binding sites to prevent actin from binding to myosin. The troponin-tropomyosin complex uses calcium ion binding to TnC to regulate when the myosin heads form cross-bridges to the actin filaments. Cross-bridge formation and filament sliding will occur when calcium is present, and the signaling process leading to calcium release and muscle contraction is known as Excitation-Contraction Coupling, which will be discussed in the next chapter.
The Neuromuscular Junction
The process of muscle contraction begins at the site where a motor neuron’s terminal meets the muscle fiber—called the neuromuscular junction (NMJ, Figure 33.6). Every skeletal muscle fiber in every skeletal muscle is innervated by a motor neuron at a NMJ. Excitation signals from the motor neuron are the only way to functionally activate skeletal muscle fibers to contract.
What Happens Next? Once the action potential travels down the sarcolemma and into the T-tubules, it triggers calcium release from the sarcoplasmic reticulum. This begins a process called excitation-contraction coupling, which leads to sarcomere shortening and muscle contraction.
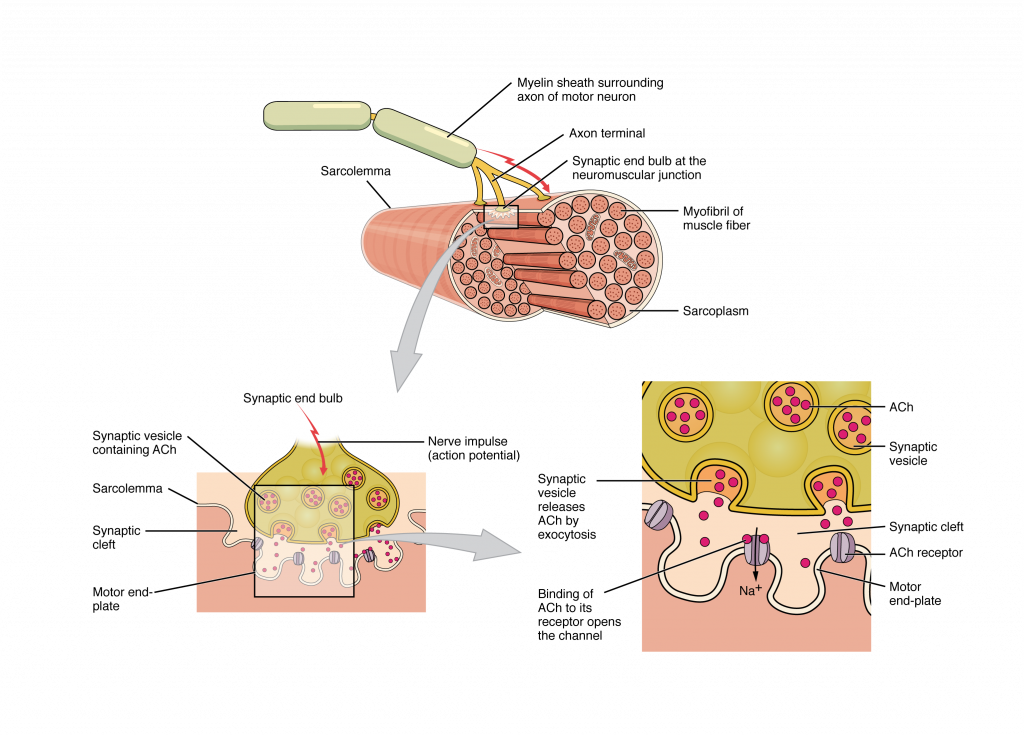
Adapted from Anatomy & Physiology by Lindsay M. Biga et al, shared under a Creative Commons Attribution-ShareAlike 4.0 International License, chapter 10.
a property of a muscle cell that enables it to respond to stimuli with changes in its membrane potential by movement of ions
a change in membrane potential that occurs due to the opening and closing of voltage-gated ion channels on the cell membrane
the ability of all muscle cells to shorten and generate force
the ability of muscle cells to stretch and extend beyond the resting length of the cell
the ability of a muscle cell to recoil to its original length after being stretched
a contractile protein; also known as the thin filament
a contractile protein; also known as the thick filament
an alternating pattern of dark and light bands of muscle myofibrils
multinucleate elongated cells with elastic sarcolemmas; also known as muscle fibers
having more than one nucleus
striated and branched muscle cells that are mononucleate, located in the heart only
non-striated involuntary muscle cells found in the walls of organs
the outermost layer of connective tissue surrounding a muscle; a sheath of dense irregular connective tissue that allows the muscle to contract while maintaining its structural integrity and separating the muscle from other tissues and organs in the area to allow the muscle to move independently
bundles of skeletal muscle fibers within a skeletal muscle
the middle layer of connective tissue surrounding a muscle; a sheath of dense irregular connective tissue that forms and surrounds fascicles
the innermost layer of connective tissue surrounding a muscle; a layer of areolar connective tissue that surrounds individual muscle fibers
a thin layer of connective tissue that connect a muscle to bone or fascia
the smooth endoplasmic reticulum of muscle fibers; the organelle that stores, releases, and retrieves calcium ions in muscle fibers
the plasma membrane of a muscle fiber
the cytoplasm of a muscle fiber
long, rod-like organelles in muscle fibers that contain myofilaments
the functional contractile unit of the muscle fiber, running from Z disc to Z disc
also known as the dark band; the band of a sarcomere containing overlapping thick and thin myofilaments
also known as the light band; the band of the sarcomere containing thin filaments only
a lighter, central region in the middle of an A band in a sarcomere, containing only thick myofilaments
the site at which a motor neuron's terminal meets a muscle fiber
